Chia sẻ:



Mô tả, chức năng và sự khác biệt của Anticodon với Codon
Một anticodon là một chuỗi gồm ba nucleotide có trong phân tử RNA chuyển (tRNA), có chức năng nhận biết một chuỗi ba nucleotide khác có trong phân tử RNA thông tin (mRNA).
Sự công nhận này giữa codon và anticodon là phản song song; nghĩa là, một cái nằm ở hướng 5 '-> 3' trong khi cái còn lại ở hướng 3 '-> 5'. Sự công nhận này giữa các chuỗi ba nucleotide (bộ ba) là nền tảng cho quá trình dịch mã; đó là, trong quá trình tổng hợp protein trong ribosome.
Do đó, trong quá trình dịch mã, các phân tử RNA thông tin được "đọc" thông qua sự nhận biết các codon của chúng bởi các anticodon của các RNA chuyển. Những phân tử này được gọi như vậy bởi vì chúng chuyển một axit amin cụ thể đến phân tử protein đang hình thành trong ribosome.
Có 20 axit amin, mỗi axit được mã hóa bởi một bộ ba cụ thể. Tuy nhiên, một số axit amin được mã hóa bởi nhiều hơn một bộ ba.
Ngoài ra, một số codon được anticodon nhận ra trong các phân tử RNA chuyển không có axit amin kèm theo; đây là những cái gọi là codon dừng.
Chỉ số
- 1 Mô tả
- 2 chức năng
- 3 Sự khác biệt giữa anticodon và codon
- 4 Giả thuyết về cán
- 4.1 RNA và axit amin
- 5 tài liệu tham khảo
Mô tả
Một anticodon được hình thành bởi một chuỗi gồm ba nucleotide có thể chứa bất kỳ bazơ nitơ nào sau đây: adenine (A), guanine (G), uracil (U) hoặc cytosine (C) trong sự kết hợp của ba nucleotide, theo cách đó nó hoạt động như một mã.
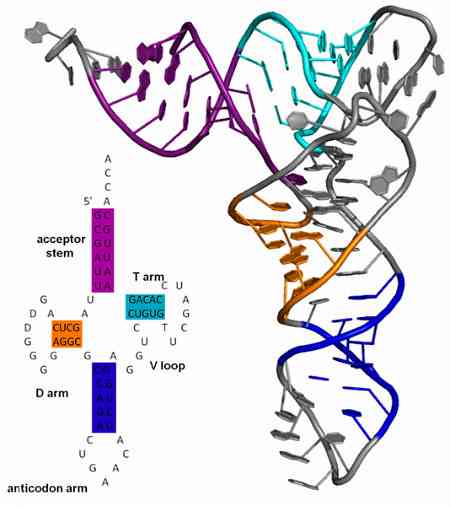
Các anticodon luôn được tìm thấy trong các phân tử RNA chuyển và luôn nằm theo hướng 3 '-> 5'. Cấu trúc của các tRNA này tương tự như cỏ ba lá, theo cách mà nó được chia thành bốn vòng (hoặc vòng); trong một trong các vòng lặp là anticodon.
Các anticodon rất cần thiết cho sự nhận biết các codon của RNA thông tin và do đó, cho quá trình tổng hợp protein trong tất cả các tế bào sống.
Chức năng
Chức năng chính của anticodon là sự nhận biết cụ thể của các bộ ba tạo thành các codon trong các phân tử RNA truyền tin. Các codon này là các hướng dẫn đã được sao chép từ một phân tử DNA để ra lệnh cho các axit amin trong protein.
Do phiên mã (tổng hợp các bản sao RNA thông tin) xảy ra theo hướng 5 '-> 3', các codon trong RNA thông tin có hướng này. Do đó, các anticodon có trong các phân tử RNA chuyển phải có hướng ngược lại, 3 '-> 5'.
Sự kết hợp này là do sự bổ sung. Ví dụ: nếu một codon là 5'-AGG-3 ', thì anticodon là 3'-UCC-5'. Kiểu tương tác cụ thể giữa codon và anticodon là một bước quan trọng cho phép trình tự nucleotide trong RNA thông tin mã hóa một chuỗi các axit amin trong protein..
Sự khác biệt giữa anticodon và codon
- Các anticodon là các đơn vị trinucleotide trong tRNA, bổ sung cho các codon trong mRNA. Chúng cho phép tRNA cung cấp các axit amin chính xác trong quá trình sản xuất protein. Ngược lại, codon là đơn vị của trinucleotide trong DNA hoặc mRNA, mã hóa một axit amin cụ thể trong tổng hợp protein.
- Các anticodon là liên kết giữa trình tự nucleotide của mRNA và trình tự axit amin của protein. Ngược lại, các codon chuyển thông tin di truyền từ hạt nhân nơi DNA đến các ribosome nơi diễn ra quá trình tổng hợp protein..
- Anticodon được tìm thấy trong nhánh Anticodon của phân tử tRNA, không giống như các codon, nằm trong phân tử DNA và mRNA.
- Anticodon là bổ sung cho codon tương ứng. Ngược lại, codon trong mRNA bổ sung cho bộ ba nucleotide của một gen nhất định trong DNA.
- Một tRNA chứa một anticodon. Ngược lại, một mRNA chứa một số codon.
Giả thuyết lăn
Giả thuyết cân bằng đề xuất rằng các điểm nối giữa nucleotide thứ ba của codon của RNA thông tin và nucleotide đầu tiên của anticodon của RNA chuyển là ít đặc hiệu hơn so với mối nối giữa hai nucleotide khác của bộ ba.
Crick đã mô tả hiện tượng này như một "rung chuyển" ở vị trí thứ ba của mỗi codon. Một cái gì đó xảy ra ở vị trí đó cho phép các công đoàn ít nghiêm ngặt hơn bình thường. Nó còn được gọi là lắc lư hoặc tamboleo.
Giả thuyết lắc lư của Crick này giải thích làm thế nào anticodon của một tRNA nhất định có thể được ghép nối với hai hoặc ba codon mRNA khác nhau.
Crick đề xuất rằng, vì việc ghép cặp cơ sở (giữa cơ sở 59 của anticodon trong tRNA và cơ sở 39 của codon trong mRNA) ít nghiêm ngặt hơn bình thường, nên cho phép "lắc lư" hoặc giảm ái lực nhất định trong trang web này.
Kết quả là, một tRNA đơn lẻ thường nhận ra hai hoặc ba trong số các codon liên quan chỉ định một axit amin nhất định.
Thông thường, các liên kết hydro giữa các bazơ của anticodon tRNA và các codon mRNA tuân theo các quy tắc nghiêm ngặt về ghép cặp cơ sở chỉ cho hai cơ sở đầu tiên của codon. Tuy nhiên, hiệu ứng này không xảy ra ở tất cả các vị trí thứ ba của tất cả các codon mRNA.
RNA và axit amin
Dựa trên giả thuyết lung lay, sự tồn tại của ít nhất hai RNA chuyển cho mỗi axit amin với codon biểu hiện thoái hóa hoàn toàn đã được dự đoán, điều này đã được chứng minh là đúng.
Giả thuyết này cũng dự đoán sự xuất hiện của ba RNA chuyển cho tất cả sáu codon serine. Thật vậy, ba tRNA cho serine đã được đặc trưng:
- TRNA cho serine 1 (anticodon AGG) liên kết với codon UCU và UCC.
- TRNA cho serine 2 (anticodon AGU) liên kết với codon UCA và UCG.
- TRNA cho serine 3 (anticodon UCG) liên kết với codon AGU và AGC.
Những đặc điểm này đã được xác minh bằng sự gắn kết kích thích của trinucleotide tinh khiết aminoacyl-tRNA với ribosome trong ống nghiệm.
Cuối cùng, một số RNA chuyển có chứa cơ sở inosine, được tạo ra từ purox hypoxanthine. Inosine được sản xuất bởi một sửa đổi sau phiên mã của adenosine.
Giả thuyết lắc lư của Crick dự đoán rằng, khi inosine xuất hiện ở đầu 5 'của anticodon (vị trí dao động), nó sẽ ghép với uracil, cytosine hoặc adenine ở codon.
Trên thực tế, alanyl-tRNA tinh khiết có chứa inosine (I) ở vị trí 5 'của anticodon liên kết với các ribosome được kích hoạt với trinucleotide của GCU, GCC hoặc GCA.
Kết quả tương tự đã thu được với các tRNA khác được tinh chế bằng inosine ở vị trí 5 'của anticodon. Do đó, giả thuyết lung lay của Crick giải thích rất rõ mối quan hệ giữa tRNA và codon được đưa ra mã di truyền, bị thoái hóa nhưng được ra lệnh.
Tài liệu tham khảo
- Brooker, R. (2012). Khái niệm về di truyền học (Tái bản lần 1). Công ty McGraw-Hill, Inc.
- Brown, T. (2006). Bộ gen 3 (3lần thứ). Khoa học vòng hoa.
- Griffiths, A., Wessler, S., Carroll, S. & Doebley, J. (2015). Giới thiệu về phân tích di truyền (Tái bản lần thứ 11). W.H. Người tự do
- Lewis, R. (2015). Di truyền học của con người: Khái niệm và ứng dụng(Tái bản lần thứ 11). Giáo dục McGraw-Hill.
- Snustad, D. & Simmons, M. (2011). Nguyên tắc di truyền(Tái bản lần thứ 6). John Wiley và con trai.