Chia sẻ:



Đặc điểm, chức năng, loại và chất ức chế Topoisomera
các topoisomeraes là một loại enzyme isomerase làm thay đổi cấu trúc liên kết của axit deoxyribonucleic (DNA), tạo ra cả sự thoải mái của nó như cuộn dây và siêu lạnh của nó.
Các enzyme này có vai trò cụ thể trong việc làm giảm căng thẳng xoắn trong DNA để các quá trình quan trọng như sao chép, sao chép DNA trong axit ribonucleic messenger (mRNA) và tái tổ hợp DNA có thể xảy ra..

Enzym Topoisomerase có mặt trong cả tế bào nhân chuẩn và tế bào nhân sơ. Sự tồn tại của nó được các nhà khoa học Watson và Crick dự đoán, khi đánh giá các giới hạn mà cấu trúc DNA đưa ra để cho phép truy cập vào thông tin của nó (được lưu trữ trong chuỗi nucleotide của nó).
Để hiểu các chức năng của topoisomera, phải xem xét rằng DNA có cấu trúc xoắn kép ổn định, với các chuỗi của nó cuộn qua nhau.
Các chuỗi tuyến tính này được cấu thành bởi 2-deoxyribose được nối bởi các liên kết phosphodiester 5'-3 'và các bazơ nitơ trong nội thất của chúng, giống như các bước của cầu thang xoắn ốc.

Nghiên cứu tô pô của các phân tử DNA đã chỉ ra rằng chúng có thể đảm nhận một số sự phù hợp tùy thuộc vào lực căng xoắn của chúng: từ trạng thái thư giãn, đến trạng thái cuộn dây khác nhau cho phép nén của chúng.
Các phân tử DNA với sự phù hợp khác nhau được gọi là topoisomers. Do đó, chúng ta có thể kết luận rằng các topoisomera I và II, có thể làm tăng hoặc giảm sức căng xoắn của các phân tử DNA, tạo thành các topoisome khác nhau của chúng.
Trong số các topoisomers DNA có thể, cấu trúc phổ biến nhất là siêu tới hạn, rất nhỏ gọn. Tuy nhiên, chuỗi xoắn kép của DNA cũng phải được giải mã bởi các topoisomera trong một số quy trình phân tử.
Chỉ số
- 1 Đặc điểm
- 1.1 Cơ chế hoạt động chung
- 1.2 Topoisomera và chu kỳ tế bào
- 2 chức năng
- 2.1 Lưu trữ nhỏ gọn vật liệu di truyền
- 2.2 Truy cập thông tin di truyền
- 2.3 Điều hòa biểu hiện gen
- 2.4 Đặc điểm của topoisomerase II
- 3 loại topoisomeraes
- 3.1-Topoisomera loại I
- 3.2 -Các loại máy ảnh loại II
- 3.3 - Topoisomeraes con người
- 4 chất ức chế Topoisomerase
- 4.1-Topoisomera là mục tiêu cho cuộc tấn công hóa học
- 4.2-Các loại ức chế
- 4.3 - Thuốc ức chế Topoisomerase
- 5 tài liệu tham khảo
Tính năng
Cơ chế hoạt động chung
Một số topoisomera chỉ có thể thư giãn các siêu tụ DNA âm tính hoặc cả hai siêu tụ DNA: dương tính và âm tính.
Nếu DNA sợi kép hình tròn không có trục trên trục dọc của nó và xảy ra rẽ trái (theo chiều kim đồng hồ), nó được cho là bị siêu âm. Nếu lần lượt là theo chiều kim đồng hồ (ngược chiều kim đồng hồ), nó cực kỳ tích cực.

Về cơ bản, topoisomeraes có thể:
-Tạo điều kiện cho sự đi qua của một chuỗi DNA thông qua một vết cắt ở chuỗi đối diện (topoisomerase loại I).
-Tạo điều kiện cho sự đi qua của một chuỗi xoắn kép hoàn chỉnh thông qua sự phân tách trong chính nó hoặc thông qua sự phân tách trong một chuỗi xoắn kép khác (topoisomerase type II).
Tóm lại, topoisomera hoạt động thông qua sự phân tách các liên kết phosphodiester, trong một hoặc cả hai sợi tạo nên DNA. Sau đó sửa đổi trạng thái cuộn dây của một chuỗi xoắn kép (topoisomerase I) hoặc hai xoắn kép (topoisomerase II), để cuối cùng quay trở lại để buộc hoặc buộc các đầu bị cắt bỏ.
Topoisomeraes và chu kỳ tế bào
Mặc dù topoisomerase I là một enzyme thể hiện hoạt động lớn hơn trong pha S (tổng hợp DNA), nó không được coi là phụ thuộc vào một giai đoạn của chu kỳ tế bào.
Trong khi hoạt động của topoisomerase II hoạt động mạnh hơn trong giai đoạn logarit của sự phát triển tế bào và trong các tế bào của các khối u đang phát triển nhanh chóng.
Chức năng
Sự thay đổi của các gen mã hóa các topoisomera gây tử vong cho các tế bào, chứng tỏ tầm quan trọng của các enzyme này. Trong số các quá trình mà topoisomera tham gia, là:
Lưu trữ nhỏ gọn của vật liệu di truyền
Topoisomera tạo điều kiện cho việc lưu trữ thông tin di truyền một cách gọn nhẹ, bởi vì chúng tạo ra cuộn DNA và siêu lạnh, cho phép tìm thấy một lượng lớn thông tin trong một khối lượng tương đối nhỏ.
Truy cập thông tin di truyền
Nếu không có topoisomera và đặc điểm độc đáo của chúng, thì không thể truy cập thông tin được lưu trữ trong DNA. Điều này là do các topoisomera định kỳ giải phóng sự căng thẳng bằng cách xoắn được tạo ra trong chuỗi xoắn kép của DNA, trong quá trình tháo gỡ của nó, trong các quá trình sao chép, sao chép và tái tổ hợp.

Nếu sự căng thẳng do xoắn được tạo ra trong các quá trình này không được giải phóng, nó có thể tạo ra một biểu hiện gen bị lỗi, sự gián đoạn của DNA tròn hoặc nhiễm sắc thể, thậm chí gây chết tế bào.
Điều hòa biểu hiện gen
Những thay đổi về hình dạng (trong cấu trúc ba chiều) của phân tử DNA làm lộ ra các vùng cụ thể ở bên ngoài, có thể tương tác với các protein liên kết DNA. Những protein này có chức năng điều hòa biểu hiện gen (tích cực hoặc tiêu cực).
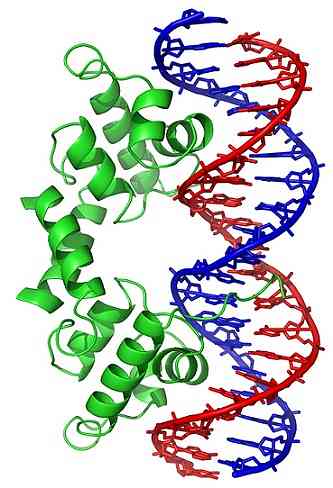
Do đó, trạng thái của cuộn dây DNA, được tạo ra bởi hoạt động của topoisomera, ảnh hưởng đến sự điều hòa biểu hiện gen.
Đặc điểm của topoisomerase II
Topoisomerase II là cần thiết cho việc lắp ráp các nhiễm sắc thể, ngưng tụ và khử nhiễm sắc thể và phân tách các phân tử DNA con gái trong quá trình nguyên phân..
Enzyme này cũng là một protein cấu trúc và là một trong những thành phần chính của ma trận của nhân tế bào trong quá trình xen kẽ.
Các loại topoisomeraes
Có hai loại topoisomera chính tùy thuộc vào việc chúng có thể phân tách một hoặc hai chuỗi DNA hay không.
-Topoisomeraes loại I
Đơn sắc
Topoisomera loại I là các monome làm giảm bớt các siêu tụ âm và dương, được tạo ra bởi sự chuyển động của ngã ba trong quá trình phiên mã và trong quá trình sao chép và tái tổ hợp gen.
Topoisomera loại I có thể được chia thành loại 1A và loại 1B. Loại thứ hai là những thứ được tìm thấy ở người và chịu trách nhiệm thư giãn DNA siêu tải.
Tyrosine trong trang web hoạt động của nó
Topoisomerase 1B (Top1B) bao gồm 765 axit amin được chia thành 4 miền cụ thể. Một trong những miền này có một khu vực được bảo tồn cao có chứa trang hoạt động với tyrosine (Tyr7233). Tất cả các topoisomera hiện diện trong trang web hoạt động của họ một tyrosine với vai trò cơ bản trong toàn bộ quá trình xúc tác.
Cơ chế hoạt động
Tyrosine từ vị trí hoạt động tạo thành liên kết cộng hóa trị với đầu 3'-phosphate của chuỗi DNA, cắt đứt và giữ nó gắn với enzyme, đồng thời chuyển một chuỗi DNA khác qua quá trình cắt bỏ.
Sự đi qua của chuỗi DNA khác thông qua chuỗi bị cắt bỏ đạt được nhờ vào sự biến đổi về hình dạng của enzyme, tạo ra sự mở của chuỗi xoắn kép DNA.
Sau đó topoisomerase tôi trở lại cấu trúc ban đầu của nó và liên kết lại các đầu bị cắt bỏ. Điều này xảy ra bởi một quá trình ngược lại với sự phá vỡ chuỗi DNA, tại vị trí xúc tác của enzyme. Cuối cùng, topoisomerase giải phóng chuỗi DNA.
Tốc độ thắt DNA cao hơn tốc độ cắt bỏ, đảm bảo sự ổn định của phân tử và tính toàn vẹn của bộ gen.
Tóm lại, topoisomerase loại I xúc tác:
- Sự cắt bỏ của một sợi.
- Các đoạn khác thông qua sự phân chia.
- Sự thắt của sự phân chia kết thúc.
-Topoisomeraes loại II
Diméricas
Topoisomera loại II là các enzyme dimeric, tách cả hai chuỗi DNA, do đó làm thư giãn các siêu tụ được tạo ra trong quá trình phiên mã và các quá trình tế bào khác.
Người phụ thuộc Mg++ và ATP
Những enzyme này cần magiê (Mg++) và cũng cần năng lượng đến từ sự cố của liên kết triphosphate ATP, chúng tận dụng lợi thế nhờ vào ATPase.
Hai trang web hoạt động với tyrosine
Con người topoisomeraes II rất giống với nấm men (Saccharomyces cerevisiae), được tạo thành từ hai monome (subfragments A và B). Mỗi monome có một miền ATPase và trong phần phụ, trang web hoạt động tyrosine 782 mà DNA có thể liên kết. Do đó, hai chuỗi DNA có thể được nối với topoisomerase II.
Cơ chế hoạt động
Cơ chế hoạt động của topoisomerase II giống như mô tả của topoisomerase I, xem xét rằng hai chuỗi DNA được phân tách và không chỉ một.
Trong vị trí hoạt động của topoisomerase II, một đoạn protein được ổn định (thông qua liên kết cộng hóa trị với tyrosine). xoắn kép của DNA, được gọi là "đoạn G". Đoạn này được phân tách và giữ liên kết với vị trí hoạt động bằng liên kết cộng hóa trị.
Sau đó, enzyme cho phép một đoạn DNA khác, được gọi là "đoạn T", đi qua đoạn "G" bị phân tách, nhờ vào sự thay đổi về hình dạng của enzyme, phụ thuộc vào quá trình thủy phân ATP.
Topoisomerase II liên kết hai đầu của "đoạn G" và cuối cùng phục hồi trạng thái ban đầu, giải phóng đoạn "G". Sau đó, DNA làm giảm căng thẳng xoắn, cho phép các quá trình sao chép và sao chép xảy ra.
-Con người
Bộ gen của con người có năm topoisomera: top1, top3α, top3β (thuộc loại I); và top2α, top2β (loại II). Các topoisomera người có liên quan nhất là top1 (topoisomerase type IB) và 2α (topoisomerase type II).
Thuốc ức chế topoisomerase
-Topoisomeraes là một mục tiêu cho cuộc tấn công hóa học
Do các quá trình được xúc tác bởi các topoisomera là cần thiết cho sự sống của các tế bào, các enzyme này là mục tiêu tấn công tốt để ảnh hưởng đến các tế bào ác tính. Đối với điều này, topoisomeraes được coi là quan trọng trong điều trị nhiều bệnh của con người.
Các loại thuốc tương tác với topoisomera hiện đang được nghiên cứu rộng rãi như các chất hóa trị liệu chống lại các tế bào ung thư (trong các cơ quan khác nhau của cơ thể) và các vi sinh vật gây bệnh.
-Các loại ức chế
Thuốc ức chế hoạt động Topoisomerase có thể:
- Nhập vào DNA.
- Ảnh hưởng đến enzyme topoisomerase.
- Xen kẽ vào một phân tử gần vị trí hoạt động của enzyme trong khi phức hợp DNA-topoisomerase được ổn định.
Sự ổn định của phức hợp thoáng qua được hình thành do sự liên kết của DNA với tyrosine của vị trí xúc tác của enzyme, ngăn chặn sự kết hợp của các đoạn bị cắt bỏ, có thể dẫn đến chết tế bào.
-Các chất ức chế topoisomeraes
Trong số các hợp chất ức chế topoisomera, như sau.
Kháng sinh chống ung thư
Thuốc kháng sinh được sử dụng chống ung thư, vì chúng ngăn chặn sự phát triển của các tế bào khối u, thường là bằng cách can thiệp vào DNA của chúng. Chúng thường được gọi là kháng sinh chống ung thư (chống ung thư). Actinomycin D, ví dụ, ảnh hưởng đến topoisomerase II và được sử dụng trong các khối u Wilms ở trẻ em và trong rhabdomyosarcomas.
Anthracyclines
Anthracyclines là một trong những loại thuốc chống ung thư hiệu quả nhất và phổ rộng nhất. Chúng được sử dụng trong điều trị ung thư phổi, buồng trứng, tử cung, dạ dày, bàng quang, vú, bệnh bạch cầu và u lympho. Nó được biết là ảnh hưởng đến topoisomerase II bằng cách xen kẽ trong DNA.
Anthracycline đầu tiên được phân lập từ vi khuẩn xạ khuẩn (Streptomyces peucetius) là daunorubicin. Sau đó, doxorubicin đã được tổng hợp trong phòng thí nghiệm, và epirubicin và idarubicin hiện cũng được sử dụng..
Bệnh than
Anthraquinones hoặc anthracenediones là các hợp chất có nguồn gốc từ anthracene, tương tự như anthracycline, ảnh hưởng đến hoạt động của topoisomerase II bằng cách xen kẽ trong DNA. Chúng được sử dụng cho ung thư vú di căn, ung thư hạch không Hodgkin (NHL) và bệnh bạch cầu.
Những loại thuốc này được tìm thấy trong các sắc tố của một số côn trùng, thực vật (frángula, senna, đại hoàng), địa y và nấm; cũng như trong hoelita, một khoáng chất tự nhiên. Tùy thuộc vào liều lượng của chúng, chúng có thể gây ung thư.
Trong số các hợp chất này, chúng tôi có mitoxantrone và losoxantrone tương tự của nó. Những thứ này ngăn chặn sự tăng sinh của các tế bào khối u ác tính, liên kết không thể đảo ngược với DNA.
Epidofilotoxin
Các chất độc tố, chẳng hạn như cholofilotoxin (VP-16) và teniposide (VM-26), tạo thành một phức hợp với topoisomerase II. Chúng được sử dụng chống ung thư phổi, tinh hoàn, bệnh bạch cầu, u lympho, ung thư buồng trứng, ung thư biểu mô tuyến vú và khối u nội sọ ác tính, trong số những người khác. Chúng được phân lập từ thực vật Podophyllum notatum và P. peltatum.
Tương tự Camptothecins
Campotecin là các hợp chất ức chế topoisomerase I, và trong số đó là irinotecan, topotecan và Diflomotecan.
Các hợp chất này đã được sử dụng để chống ung thư ruột kết, phổi và vú, và được lấy tự nhiên từ vỏ và lá của các loài arboreal Camptotheca acuminata đào Trung Quốc và Tây Tạng.
Ức chế tự nhiên
Sự thay đổi cấu trúc của topoisomeraes I và II cũng có thể xảy ra hoàn toàn tự nhiên. Điều này có thể xảy ra trong một số sự kiện ảnh hưởng đến quá trình xúc tác của nó.
Trong số những thay đổi này, chúng ta có thể đề cập đến sự hình thành các chất làm mờ pyrimidine, sự không phù hợp của các bazơ nitơ và các sự kiện khác gây ra bởi stress oxy hóa.
Tài liệu tham khảo
- Anderson, H., & Roberge, M. (1992). DNA topoisomerase II: Một đánh giá về sự tham gia của nó vào cấu trúc nhiễm sắc thể, sao chép DNA, phiên mã và nguyên phân. Báo cáo quốc tế về sinh học tế bào, 16 (8): 717-724. doi: 10.1016 / s0309-1651 (05) 80016-5
- Chhatriwala, H., Jafri, N., & Salgia, R. (2006). Một đánh giá về ức chế topoisomerase trong ung thư phổi. Sinh học & Trị liệu Ung thư, 5 (12): 1600-1607. doi: 10,4161 / cbt.5.12,3546
- Hồ, Y.-P., Au-Yeung, S.C. F., & To, K. K.W. (2003). Tác nhân chống ung thư dựa trên bạch kim: Chiến lược thiết kế sáng tạo và quan điểm sinh học. Nhận xét nghiên cứu y học, 23 (5): 633-655. doi: 10.1002 / med.10038
- Li, T.-K., & Liu, L. F. (2001). Sự chết của tế bào khối u gây ra bởi các thuốc nhắm mục tiêu topoisomerase. Đánh giá hàng năm về Dược lý và Độc tính, 41 (1): 53-77. doi: 10.1146 / annurev.pharmtox.41.1.53
- Lưu, L. F. (1994). DNA Topoisomeraes: Thuốc nhắm mục tiêu Topoisomerase. Báo chí học thuật. trang 307
- Osheroff, N. và Bjornsti, M. (2001). DNA Topoisomerase. Enzymology và thuốc. Tập II. Báo chí Humana. trang 329.
- Rothnberg, M. L. (1997). Các chất ức chế Topoisomerase I: Xem lại và cập nhật. Biên niên sử về ung thư, 8 (9), 837-855. doi: 10.1023 / a: 1008270717294
- Ryan B. (2009, ngày 14 tháng 12). Topoisomerase 1 và 2. [Tệp video]. Lấy từ youtube.com